Induction of the heat shock protein in rat lungs following
intermittent hypoxic training
KoDo Okuyama 1), Jingtao Jiang 2)
Clinical Research Laboratory of 'Mountain Air' Therapy 2-21-15
Shimouma Setagaya-ku Tokyo JAPAN
Central Institute for Electron Microscopic Researches, Nippon
Medical School, Sendagi, Bunkyo-ku, Tokyo, Japan.
Abstract
In order to demonstrate the mechanism of intermittent hypoxic
training (IHT), we studied the expression of HSP70 by immunohistochemistry
using the Streptavidin Biotin Peroxidase Complex(SABC) Method
on alveolar type I , type II epithelial cells, macrophages
from rats with IHT. The expression of HSP70 in IHT in comparison
with control was significantly increased on alveolar type
I , type II epithelial cells, macrophages and was significantly
correlated with the duration of IHT.
This study demonstrates that expression of HSP70 may be a mechanism
of adaptation of hypoxia by IHT.
Key words: Intermittent hypoxic training HSP70 Electron microcopy Immunohistochemistry
lntroduction
Intermittent hypoxic training (IHT), with repeated short-term
inhalation of hypoxic mixtures, has been used to treat and
prevent certain diseases and has a very favorable effect for
the exercise of athletes 1,2. Investigations of IHT have showed
that increased hypoxic ventilatory response (HVR) 3,4 is an
important physiological response. Resent studies have displayed
that IHT inhibits the free radical production, which gives
harmful effect to the cells and tissues, and raise the metabolic
rate as a result of sympathetic nervous system activation
5,6. Rats trained to intermittent normobaric hypoxia developed
an increase of the glycogen contents in the heart and liver
parenchymatous cells and offered as many beneficial effects
in protecting against myocardial injuries 7,8. Antixidant
enzymes and stress proteins may be part of the mechanisms
contributing to the cardioprotection of the intermittent hypoxic
adaptation 7.Heat shock causes intracellular expression of
a specific group of proteins called heat steins (HSPs) that
have broad cytoprotective properties 9,10. The first demonstration
of HSP- mediated cytoprotection involved the phenomenon of
thermotolerance, whereby a brief heat shock conferred protectionagainst
subsequent exposure to otherwise lethal hyperthermia 11. Subsequent
studies demonstrated that induction of HSPs also protected
cells and whole organs against nonthermal cytotoxic agents
such as oxidants, nitric oxide, tumor necrosis factor-α, and
endotoxin 12-14. Previous studies also demonstrated that induction
of HSPs protected against in vitro and in vivo models of acute
lung injury and may have therapeutic value for attenuating
acute lung injury . HSP 70 has been shown to be protective
following ischemic injury. Similarly, a member of the small
heat shock family, HSP27 has been shown to play a role in
cellular repair and mechanisms of protection against cell
stress. In this study, we are used monoclonal antibody to
heat shock protein 70 to investigate whether IHT induces the
expression of HSP70 in lungs.
Materials and methods
1. Animals
A total of 18 male Wistar rats, aged 12 weeks were used: three
animals for control and rest for the experimental groups.
They were allowed free access to food and water during intermittent
hypoxic training.
2.Intermittent hypoxic training:
Hypoxia was induced by exposure to 10% oxygen with machine.
The rats were sustained hypoxia for 15min one time a day for
3, 7, or 21 consecutive days, respectively. Control rats exposured
toroom air.
3. Light and electron microscopy.
At 3D, 7D and 21 Days after intermittent hypoxic training
(IHT), the animals were anesthetized with pentobarbital sodium
(50mg/kg) and lungs were removed. For light microscopic examination,
the tissues were fixed with 10% formalin in phosphate buffer
solution, and embedded in paraffin.
Then paraffin sections of 2 μm in thickness were prepared
and stained with hematoxylin and eosin. For electron microscopic
examination, rat lung specimens were cut into 2mm3 blocks,
fixed 2.5% glutaraldehyde in 0.1M phosphate buffer and
postfixed with 1% osmium tetroxide, dehydrated in a graded
alcohol series and embedded in Epon 812. Semithin sections
stained with toluidine blue were used for high light microscopy
and selection of areas for thin sectioning. Thin sections
were cut with 5000 Ultrotome, stained with uranyl acetate
and lead citrate. The sections were examined under a JEM-
1010 transmission electron microscope.
4. Immunohistochemistry
The Streptavidin Biotin Peroxidase Complex (SABC) Method was employed
for immunohistochemistry for heat shock protein (HSF70). Briefly,
deparaffinized sections were treated for 30 min with 0.3%
H2O2 to block endogenous peroxidase and rinsed again 3×5 min
inPBS. Sections were then incubated for 30 min with block
nonspecific reactive sites by applying 1 :20 normal goat serum
and incubated with Monoclonal anti-HSP antibody (NOVO) diluted
1:40 in PBS for 60 min. After washing in PBS, they were incubated
with goat anti-mouse immunoglobulins for 10 min and then incubated
with a mixture of streptavidin and biotinylated horseradish peroxidase
for 5 min. After washing in PBS, reacted with 0.05% 3 - 3'
diaminobenzidine (DAB) containing 0.01 %H2O2.
Results
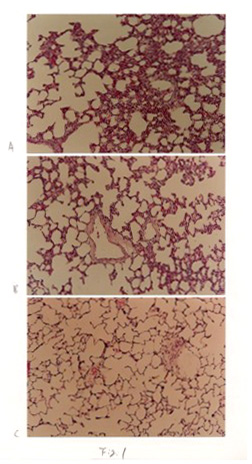
Light microscopic observations
The morphological observations in the control group were normal
from 3 days to 21 days. After three days IHT, the remarkable
interstitial edema,increased thickness of the alveolar septa,
marked capillary dilatation,proliferation of interstitial
cells, the collapse of alveoli and dilation of the alveolar
ducts were observed compared with controls (Fig. 1A). After
7 days IHT, those pathologic changes decreased than 3 days
(Fig.1B). After 21 days of IHT, the nearly normal structure
of lungs was observed (Fig.1C).
Immunohistochemical observation
The heat shock protein 70 monoclonal antibody was used to
stain the lung tissue from the control and IHT rats immunohistochemically.
In control lungs, there are weak positive staining for HSP70
observed in the bronchial epithelial cells as well as in some
alveolar type II epithelial cells.
After 3 days IHT, HSP70 was moderately expressed in alveolar
type I ,type II epithelial cells, macrophages and bronchial
epithelial cells. HSP70 expressed in cytoplasm and nuclei
(Fig.2A). After 7 days IHT, expression of HSP70 was same as
3 days IHT (Fig.2B). After 21 days IHT, there was strong septa
also were displayed. By 7 days, the congestion of capillary,
type II cell proliferation, and lipid drops in alveolar septa
were observed. By 21 days,congestion of capillary, increased
capillaryendothelial volume, lipid drops in the alveolar septa
and proliferation of alveolar type II cells were noted(Fig.3B).
Discussion :
The principle of intermittent hypoxic training (IHT), with
repeated short-term inhalation of hypoxic mixtures had been
proposed by S. Strelkov and his associates in the early of
1980's based on their obstetrical practice. Intermittent hypoxic
training (IHT) has showed promise for prevention and treatment
of some diseases and efficiently produces great advancement
in athletic training 1,2. The mechanism of IHT remains unidentified.
A number of mechanisms have been postulated including optimizing
both hypothalamic-pituitary-adrenal axis functioning and free
radical-mediated process control 5, increase of the quantity
and secretory activity of peptidergic neurons of the paraventricular
hypothalamic nucleus (PHN) 16,enhancement of neurotransmission
in the carotid body (CB) as well as in central
structures through NADPH oxidase stimulation 17, increase
in ventilatory response under repetitive hypoxia, changes
in suprapontine facilitation of resporatory activity and Changes
in monoamine metabolism or release 18, and raise the metabolic
rate as a result of sympathetic nervous system activation
6.
In our clinical practice, we found that IHT could release
the stress in some patients. In this study, we found that
IHT induced the expression of HSP70. The response of cells
or organisms to stress such as exposure to heat or chemicals
is associated with the induction of heat shock proteins (HSPs).
Heat shock proteins (HSPs) are an evolutionarily conserved
group of proteins that are highly inducible by a wide variety
of stressors. HSPs are grouped by molecular weight and amino
acid sequence similarity into five main families: The high
molecular weight 100-110kDa family; the 83-90kDa family; the
70kDa family ranging from 66 to 78kDa and containing the highly
inducible HSP 70; the 60kDa family present in bacteria,mitochondria,
and chloroplasts; and a diverse group of small HSPs ranging
from 15 to 30kDa. Of great interest are observations demonstrating
that once a heat shock response has been induced, the cells
or organs can show remarkable resistance to subsequent metabolic
stress. Heat shock protein 70 (HSP70) has been shown to have
a protective role in ischemic disease,inflammation, infection
and a potential role in antigen processing as well as a possible
regulatory role in cytokine biosythesis 13,14. HSP70 exists
in the cell in equilibrium between its free state, in the
cytoplasm, and its bound state, protecting proteins in the
nucleolus, perhaps either by helping refold some of the unfolded
ribosomal proteins or by solubilising the denatured ribosomal
proteins to facilitate their turnover. During release from
heat shock and as the nucleoli begin to recover their normal
activities, most of the HSP 70 returns to the cytoplasm. Stress
proteins have an important role in normal cellular physiology
apart from participation in the stress response. Under normal
conditions, stress proteins are involved in the successful
folding, assembly, intracellular localization, and secretion
of nascent protein chains as they emerge from the ribosome.
Stress proteins also function to regulate the degradation
of proteins to prevent the accumulation of protein aggregates
within the cell. Cultured bovien and ovine pulmonory artery
endothelial cells and guinea pig airway epithelial cells and
alveolary macrophages expressed abundant HSP 70 after thermal
stress14, 19, 20. In rabbit alveolar type II cells, the process
of cell isolation itself induced stress protein expression
21. In vivo thermal stress increased stress protein expression
in the lungs and other organs of rats, and the time courses
and relative magnitudes of expression differed among organs
22,23. Bonay et al demonstrated limited stress protein expression
in normal human lungs. HSP90, HSP 70 and HSP63 were selectively
expressed in proximal bronchiolar epithelium and alveolar
macrophage. In contrast, more distal bronchiolar epithelium,
type I and type II alveolar cells, and stromal cells did not
express stress proteins 24. HSP 70 expression was substantially
increased in airway epithelium and alveolar macrophages of
patients with asthma compared with control subjects 25. In
this study, we observed the morphologic changes of lungs and
expression of HSP70 after rat IHT. We found that that pulmonary
damage occurred at 3 days IHT, one week IHT later, the pulmonary
damage repaired in hypoxic animals. The expression of HSP70
observed in alveolar type I, type IIepithelial cells,macrophages
and bronchial epithelial cells after 3 dr 3 days IHT, andcontinually
expressed until 2l days IHT. Expression of HSP 70 in 21 days
IHT was stronger than 3 days IHT. This revealed that IHT could
induce HSP70 in alveolar type I , type II epithelial cells,
macrophages and bronchial epithelial cellsveolar type I, type
II epithelial cells, macrophages and bronchial epithelial
cells after IHT. HSP70 expressed after 3 days IHT, and continued
to 21 days IHT. Although the exact significance of these data
is still unresolved, it is proposed expression of HSP may
be a mechanism of adaptation of hypoxia with IHT.
Reference
1 Serebrovskaya T, Swanson R, Karaban IN, Serebrovskaya Z,
Kolesnikova EE. Intermittent hypoxia alters hypoxic ventilatory
responses. Fiziol Zh. 45(5): 9-18, l999
2 Serebrovskaya T, Swarotective effect of stress protein induction
in a rat model of acute lung injury caused by intratracheal
administration of phospholipase A1 and systemic administration
of endotoxin 22,23.This study is the first to demonstrate
an expression of HSP 70 in abbins PA.Alterations in respiratory
control during 8 h of isocapnic and poikilocapnic hypoxia
in humans. J Appl Physiol. 78: 1098-107,1995
4 Schoene RB, Roach RC, Hackett PH, Sutton JR, Cymerman A,
Houston CS, Operation Everest II: ventilatory adaptation during
gradual decompression to extreme altitude. Med Sci Sports
& Exercise. 22:804- 10,1990
5 Adiiatulin AI, Piliavskaia AN, Takchuk EN, Guliaeva NV,
[Various mechanisms of protective action of interval hypoxic
training during preparation for abdominal dapy. Adaptation
Biology and Medicine(Vol.3).
6 Cao KY, Zwillich CW, Berthon-Jones M, Sullivan CE. Increased normoxic
ventilation induced by repetitive hypoxia inconscious dogs.
J Appl Physiol. 73:2083.8, l992
7. Zhuang J, Zhou Z. Protective effects of intermittent hypoxic
adaptation on
myocardium and its mechanisms. Biol Sinals Recept. 8:316-22,1999.Review.
8. Lebkova NP, Chizhov AI, Bobkov II. The adaptational intracellular
mechanism regulating energy homeostasis during intermittent
normobaric hypoxia. Ross Fiziol Zh lm I M Sechenova. 85:403-11,
1999
9. Minowada G, Welch WJ: Clinical implications of the stress
response. J Clin. Invest. 95:3-12, 1995
10. Wang HR, Eispe JR: The stress response and the lung. Am
J Physiol 273: L1-9, l997
11. Gerner EW, Schneider MJ: Induced thermal resistance in
Hela cells. Nature 256:500-2, 1975
12. Meerson FZ, Malyshev I Yu, Zamotrinsky AV. Differences
in adaptive stabilization of structures in response to stress
and hypoxia relate with the accumulation of hsp 70 isoforms.
Mol Cell Biochem. 111:87-95, 1992
13. Koh Y, Lim CM, Kim MJ, Shin TS, Lee SD, Kim WS, Kim DS,
Kim WD. Heat shock response decrease endotoxin-induced acute
lung injury in rats. Respirology.4:325-30, 1999
14. Wong HR, R.J Mannix, J.M. Rusnak, A. Boota, H. Zar, S.
C. Watkins, J.S. Lazo,B.R. Pitt. The heat shock response attenuates
lipopolysaccharide-mediated apoptosis in cultured sheep pulmonary
artery endothelial cells. Am. J. Respir. Cell Mol. Biol. :745-51,
1996
15. Wong HR, Ryan M, Menedez IY, Denenberg A and Wispe JRlar
hypothalamic nucleus and brain stem neurons in rats]. [Russian]
Ross Fiziol Zh lm I. M. Sechenova. 84(3): 173-81, 1998
17 0lano M, Song D, Murphy S, Wilson DF, Pastuszko A. Relationships
of dopamine, cortical oxygen pressure, and hydroxyl radicals
in brain of newborn piglets during hypoxia and posthypoxic
recovery. J Neurochem. 65(3):1205-12, 1995
18. Soto-Arape I, Burton MD, Kazemi H. Central amino acid
neurotransmitters and the hypoxic ventilatory response. American
J Respir Crit Care Med. 151(4):1113-20, 1995
19. Rinaldo J. E, M. Gorry R, Stricter H, Cowan R, Abdolrasulnia
V. Shepard. Effect of endotoxin-induced cell injury on 70-kD
heat shock proteins in bovine lung endothelial cells. Am.
J. Respir. Cell Mol. Biol. 3:207-16, 1990
20. Cohen D.S, E. Palmer WJ, Welch, D. Sheppard. The respose
of guinea pig airway epithelial cells and alveolar macrophages
to environmental stress. Am. J. Respir. Cell Mol. Biol. 5:133-43,
1991
21. Brandes M. E, J. N Finkelstein. Induction of the stress
response by isolation of rabbit type II pneumocytes. Exp.
Lung.Res. 15: 93-111, 1989.
22. Villar J, J. D. Edelson, M. Post, B. Mullen, A. S. Slutsky.
Induction of heat Stress proteins is associated with decreased
mortality in an animal model of acute lung injury. Am. Rev.
Respir. Dis. l47:177-81, l993.
23. Villar J.,S.P.Ribeiro,J.B.M.Mullen,M.Kuliszewski,M.Post,A.S.Slutsky.Induction
of the heat shock response reduces mortality rate and organ
damage in a sepsis-induced acute lung injury model. Crit.
Care Med. 22:9 B ± through preventing I o B kinase activation
in 0respiratory epithelial cells. J. Immunol 164:541 6-23,
2000
27. Yoo, C.G., Lee S., Lee C. T., Rim Y. W., Ham S. K., and
Shin Y. S. Anti-inflammatory effect of heat shock protein
induction is related to stabilization of I κ B α through preventing
I κ B kinase activation in respiratory epithelial cells. J.
Immunol 164:5416-23, 2000
28. Simon M. M, A. Reikerstolfer, A. Schwarz, C. Kronis, T.
G.Lunger, M. Jaatela, T. Schwarz. Heat shock protein 70 overexpression
affects the response to ultraviolet light in murine fibroblast.
J. Clin. Invest. 95:926-933,1995
Figure Legends
Fig 1 Change of light microscope in IHT rat lungs. A: 3 days
IHT. B:7days IHT.
C: 21 days IHT.
Fig 2 Immunohistochemical staining of rat lung with anti-HSP70
after IHT.
A: 3 days IHT. B: 7 days IHT. C: 21 days IHT.
Fig 3 Ultrastructural changes of lung in IHT: A: 3 days IHT.
B: 21 days IHT
|